Mikrostrukturen im lebenden Gehirn erkennen: Magnetresonanztomografie der Myelinscheide
Forschungsbericht (importiert) 2009 - Max-Planck-Institut für Kognitions- und Neurowissenschaften
Die signalübertragenden Fortsätze (Axone) von Nervenzellen sind über einen Großteil ihrer Länge von einer mehrlagigen Membranschicht, dem Myelin, umhüllt. Quantitative Informationen über die Myelinscheide sind wichtig, um neurologische Erkrankungen, die Entwicklung des Nervensystems oder die Parzellierung der Hirnrinde in funktionell abgrenzbare Bereiche zu verstehen. Bislang beruhen die Kenntnisse hierzu weitgehend Gewebeuntersuchungen, die nach dem Tod (post mortem) durchgeführt wurden. Neue Ansätze der Magnetresonanztomografie (MRT) zielen dagegen auf eine bildgebende Darstellung der Myelinisierung am lebenden Organismus (in vivo), die ohne Eingriff von außen erreicht wird.
 mit sich wiederholenden Membrandoppelschichten (B); das dazwischen befindliche Myelinwasser wird anhand seiner kurzen longitudinalen Relaxationszeit T1 vom sonstigen Gewebewasser unterschieden (C).")
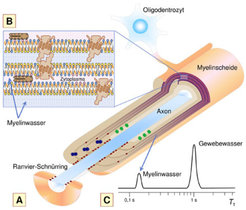
Aufbau der Myelinscheide
Die Myelinscheide wird im Gehirn von einem besonderen Zelltyp, den Oligodendrozyten, gebildet, indem diese ihre Plasmamembran in einer engen Spirale um das Axon wickeln (Abb. 1A). Die sich wiederholenden Doppelschichten der Myelinmembran bestehen zu etwa 70 Prozent aus Lipiden (Fetten) und zu etwa 30 Prozent aus Proteinen (Abb. 1B). Zwischen diesen Einheiten befindet sich Wasser, das etwa 40 Prozent des Gesamtgewichts des Myelins ausmacht („Myelinwasser“). Die Myelinscheide fungiert unter anderem als elektrischer Isolator des Axons. Sie ist in Längsrichtung in regelmäßigen Abständen von Lücken, den Ranvier-Schnürringen, unterbrochen. Durch die segmentierte Myelinanordnung wird die Nervenleitungsgeschwindigkeit stark erhöht. Einen hohen Myelinanteil besitzt das nervenfaserreiche Marklager (weiße Hirnsubstanz). In geringerem Ausmaß enthält auch die Hirnrinde (graue Hirnsubstanz) Myelin.
Magnetresonanztomografie und Relaxation
Die Magnetresonanztomografie nutzt die Ausrichtung der magnetischen Dipolmomente von Wasserstoffatomkernen in einem starken Magnetfeld. Dabei wird eine schwache, in Feldrichtung weisende Magnetisierung aufgebaut. Der Prozess des Magnetisierungsaufbaus wird auch als „longitudinale Relaxation“ (oder „Längsrelaxation“) bezeichnet und benötigt eine charakteristische Zeit, die sogenannte „longitudinale Relaxationszeit“ T1. Durch Einstrahlung eines Hochfrequenzpulses lässt sich die Magnetisierung aus der Gleichgewichtslage kippen. Somit entsteht eine Komponente quer zum externen Feld, die sich wie ein Kreisel um die Feldachse dreht. Dies erzeugt nach dem Dynamoprinzip in einer Empfangsspule ein schwaches Induktionssignal, dem zur Berechnung von Schnittbildern noch eine Ortskodierung aufgeprägt wird.
Die durch Pulseinstrahlung gestörte Magnetisierung ist bestrebt, wieder in die Gleichgewichtslage zurückzukehren. Dieser Relaxationsprozess kann durch Exponentialgesetze beschrieben werden. Einerseits läuft die bereits genannte longitudinale Relaxation ab, bei der die in Feldrichtung weisende Magnetisierungskomponente wieder aufgebaut wird. Daneben zerfällt die Querkomponente – und damit das Induktionssignal – mit einer charakteristischen „transversalen Relaxationszeit“ T2.
Bei der Magnetresonanztomografie des Gehirns tragen zum Signal nahezu ausschließlich Wassermoleküle bei, da diese in hoher Konzentration vorhanden sind und aufgrund ihrer hohen Beweglichkeit ein günstiges Relaxationsverhalten zeigen. Dagegen ist es nicht möglich, Membrankomponenten direkt abzubilden, denn ihre Signale zerfallen innerhalb von weniger als 100 Mikrosekunden [1]. Ein indirekter Ansatz, der das Verhalten des Wassers im Gewebe untersucht, gestattet es, Informationen über die Myelinscheide abzubilden (Abb. 1C). Dabei wird anhand der Relaxationseigenschaften das Wassersignal in verschiedene Komponenten aufgetrennt [2].
Relaxografische Bildgebung
Im Unterschied zur konventionellen Magnetresonanztomografie, die die Magnetisierung in einem einzigen, vorab festgelegten Relaxationszustand abbildet, wird bei der Relaxografie eine vollständige Relaxationskurve mit vielen Abtastpunkten aufgezeichnet (Abb. 2A). Für die T1-Relaxografie wird dazu zunächst die Magnetisierung umgekehrt und dann der Wiederaufbau des longitudinalen Gleichgewichtswerts durch eine Serie von Pulsen mit kleinem Kippwinkel verfolgt. Die weitere Analyse besteht darin, aus diesen als Funktion der Zeit aufgenommenen Signalen für jeden Bildpunkt die T1-Verteilung („Relaxogramm“) zu errechnen – mathematisch entspricht dies einer inversen Laplace-Transformation (ILT). Allerdings ist die ILT vom mathematischen Standpunkt her ein „schlecht gestelltes“ Problem: Ohne weitere Maßnahmen werden die Signale in den erhaltenen Relaxogramme durch Rauschen so stark verbreitert, dass keine detaillierte Aussage möglich ist. Um die Analyse zu stabilisieren, wird daher eine Regularisierung vorgenommen, bei der man nach dem „Sparsamkeitsprinzip“ von einem glatten Relaxogramm mit wenigen Komponenten ausgeht. Anders ausgedrückt bedeutet dies, dass die T1-Verteilung keine Sprünge oder Kanten besitzen soll.
 und daraus ein Relaxogramm errechnet (B, C). Bei 3 Tesla liegen die T1-Signale des Myelinwassers bei ca. 0,1 Sekunden und des Gewebewassers bei ca. 1,1 Sekunden (B). Bei 7 Tesla wandert das Gewebewassersignal nach rechts, und das Myelinwassersignal wächst, da Austauschprozesse die Messung weniger stark beeinflussen (C). Damit wird die Konzentrationsabschätzung zuverlässiger. Die Falschfarbenkarten zeigen die räumliche Myelinwasserverteilung in Prozent bezogen auf den gesamten Wassergehalt.")
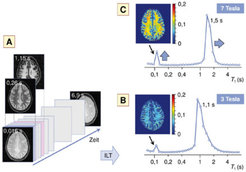
Eine Besonderheit der am Max-Planck-Institut für Kognitions- und Neurowissenschaften entwickelten Methode liegt darin, dass die Regularisierung nicht nur entlang der T1-Achse vorgenommen wird. Vielmehr nehmen die Wissenschaftler an, dass auch die räumliche Nachbarschaft der Bildpunkte eine glatte Verteilung hat; man spricht daher auch von „Kreuzregularisierung“ [3]. Damit lässt sich die Schärfe der Relaxogramme weiter verbessern, und verschiedene Wasseranteile können getrennt werden (Abb. 2B, C). Unter anderem sind dies das zelluläre und außerzelluläre Gewebewasser mit einer T1-Zeit im Sekundenbereich und eine Komponente mit kurzer T1-Zeit (etwa 100 Millisekunden), die dem Myelinwasser zugeordnet werden kann [4]. Der gefundene Myelinwasseranteil für die weiße Substanz von etwa 12 Prozent (gelbe Farbe in Abb. 2C) stimmt sehr gut mit Abschätzungen aus T2-Messungen [1] überein. Ein Vorteil der T1-Relaxografie besteht darin, dass sie mit einer geringeren Hochfrequenzleistung auskommt und so die thermische Belastung des Gewebes reduziert wird. Erwartungsgemäß ist der gefundene Myelinwasseranteil in der grauen Substanz deutlich geringer (ca. 5 Prozent; blaue Farbe in Abb. 2C).
So wie bei einer Filmkamera Bewegungen nur bei ausreichender Verschlussgeschwindigkeit scharf abgebildet werden, ist eine relaxografische Bestimmung von Myelinwasser nur möglich, wenn der Wasseraustausch zwischen Myelin und den angrenzenden Gewebebereichen nicht zu schnell erfolgt. Die Verschlussgeschwindigkeit wird im vorliegenden Fall durch den Unterschied der Relaxationsraten beider Komponenten festgelegt. Modellrechnungen und die Relaxogramme zeigen, dass der Austausch langsam genug abläuft, um Myelinwasser zu identifizieren. Für stärkere Magnetfelder werden die Messbedingungen aufgrund längerer T1-Zeiten besser (größere Verschlussgeschwindigkeit). Experimente bei 3 Tesla (Abb. 2B) zeigten daher einen scheinbar geringeren Myelinwasseranteil als solche bei 7 Tesla (Abb. 2C) [5]. Es ist zu erwarten, dass Veränderungen der Myelinmembran durch Erkrankungen (beispielsweise Multiple Sklerose) oder Alterungsprozesse die Austauschgeschwindigkeit beeinflussen. Zukünftig geplante Experimente sollen daher zeigen, ob sich aus dem T1-Relaxogramm Indikatoren für Demyelinisierungsprozesse ableiten lassen.